

Розуміння особливостей формування та адаптивного значення статевої структури популяцій тварин (в тому числі, риб) можливо тільки при розгляді в цілому теорії статі, механізмів визначення статі, а також конкретних ситуацій проживання популяцій і утворюється у них при цьому статевої структури. Необхідно розмежовувати поняття «співвідношення статей в популяції» і «статева структура популяції» (Коновалов, 1980).
Перше поняття має сенс, давно прийнятий в біологічній літературі, і визначає частку самців і самок в розмірної, вікової чи іншій групі, а також в популяції на будь-якому відрізку часу її існування.
Поняття «статева структура популяції» характеризує, з одного боку, співвідношення статей тільки в репродуктивний період, а з іншого - воно включає в якості елемента структури характеристику часткою самців і самок в кожній віковій групі, які можуть різнитися досить істотним співвідношенням статей.
У природі у живих організмів існує три основних типи розмноження. Перший тип - це безстатеве, засноване на мітотичного ділення клітин. Другий тип - хибно-статеве розмноження є різновидом безстатевого розмноження у організмів, що мають статеву систему. Третій тип - статеве розмноження пов'язане з формуванням чоловічих і жіночих статевих систем, репродуктивним розподілом статевих клітин під час мейозу і розвитку зародка тільки після злиття гамет (Коновалов, 1980).
Для хребетних відомо два останніх способу розмноження. Прикладом помилково-статевого розмноження може служити партеногенез, а у риб, зокрема, - гіногенез, при якому інактивована ядро чоловічої гамети є джерелом дезорганізованого хроматину. Останній був докладно описаний у карася.
Істинний статевий процес може відбуватися при формуванні жіночих і чоловічих статевих органів в одному організмі (гермафродитизм) або в різних: у самців і самок. Гермафродит поширений у риб і існує в декількох формах, що ми не розглядаємо, т. К. Це не є метою цієї роботи.
Знахідки гермафродитних особин у тихоокеанських лососів вкрай рідкісні. Відомо лише обмежене число фактів за вікову історію інтенсивного дослідження та експлуатації лососів (Коновалов, 1980). У всіх відомих випадках у лососевих зустрічалися одночасно насінники і яєчники (або статеві залози, що складаються частково з сім'яників і частково з яєчників). Для тихоокеанських лососів синхронний гермафродитизм в обох формах був описаний для чавичі (Rutter, 1904), кіжуча (Grawford, 1927) і кети (Uzmann, Hesselholt, 1958) - все цитованості по: Коновалов, 1980.
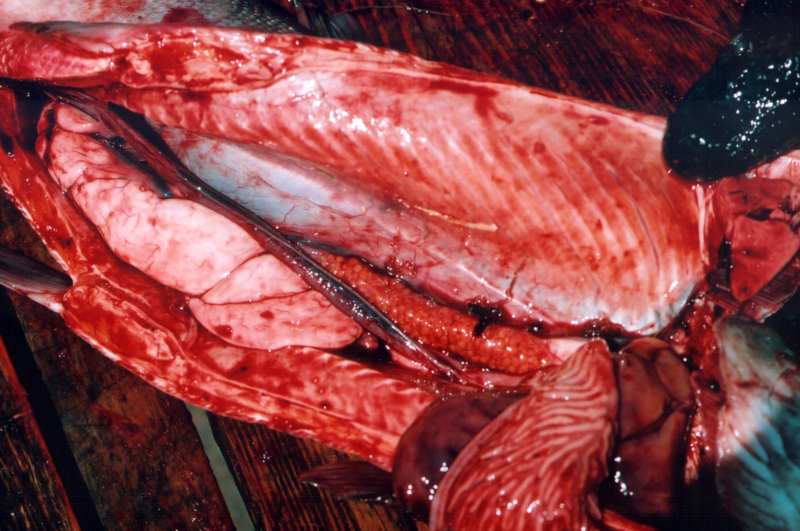
Ми не у своєму розпорядженні сучасні даними за американськими стадам, але опублікований приклад гермафродитизму у вітчизняній літературі для азіатської нерки в першій формі вже з'явився (Бугаєв, 2007). Перш, цього автору, за 40-річний період його діяльності, двічі зустрічалися гермафродити у нерки в першій формі (див. Фото) і один раз - у другій.
Р. Фішер (Fisher, 1930 - цит. За: Коновалов, 1980), розвиваючи вчення Ч. Дарвіна (Darwin, 1964) про статеве відборі, намагався докласти до нього закони успадкування. Він прийшов до висновку про перевагу статевого розмноження з генетичних позицій в зв'язку з тим, що збільшується число рекомбінацій генів і створюються умови, що сприяють підтриманню максимального генетичного різноманіття.
У свою чергу, відбір в різних частинах ареалу може істотно відрізнятися, і, таким чином, по ареалу може підтримуватися значну різноманітність завдяки статевого розмноження. Р. Фішер (Fisher, 1930) показав, що «дарвиновское» перевагу самками одних самців іншим легко можна пояснити з генетичних позицій, і воно виражається через диференційовані можливості до відтворення. Він надавав важливе значення інформації про співвідношенні статей в період розмноження, особливо в обмежених за чисельністю популяціях. Надалі висунуті Р. Фішером (Fisher, 1930) положення були розвинені іншими дослідниками.
На думку С. М. Коновалова (1980), статеве розмноження проявляє всі свої достоїнства при проживанні виду в помітно змінного навколишнього середовища. В цьому випадку безстатеве розмноження фактично не може звести до мінімуму втрати в чисельності при переході з одного адаптивного стану популяції в інше. Організми з статевим розмноженням при зміні умов проживання зменшуються в чисельності, по через "n" поколінь можуть відновити чисельність. Однак в останньому випадку домінують особини, адаптовані до знову встановилися умов навколишнього середовища.
Вся історія досліджень тихоокеанських лососів свідчить, що на розміри і масу тіла дозрілих риб (на нерестовищах - виробників) істотно впливає їх вік статевого дозрівання, пов'язаний з тривалістю перебування їх в море.
Як далі пише С. М. Коновалов (1980, стор. 145-146): «Ідея вдосконалення адаптацій виду до мінливих умов середовища шляхом закріплення статевого розмноження і подальшої диференціації статей розвинена в циклі робіт В. А. Геодакяна (1965a-b, 1971 ; Геодакян, Смирнов, 1968; Геодакян, Кособуцький, 1969). Він вважає, що до потомству існує два канали зв'язку - від самця і від самки і чим більше різноманітність самців, тим їх канал зв'язку буде мати більш широке розтин в порівнянні з каналом самок. У цьому випадку кількість самок визначає чисельність потомства, а кількість самців - швидкість зміни середнього генотипу. В. А. Геодакян доводить, що число самців має зростати при несприятливих умовах середовища, що вимагають пристосованості, при цьому повинна зростати «оборотність» самців.
Конкретизуючи, можна підкреслити, що В. А. Геодакян передбачає підвищену мінливість самців за генетичними, фізіологічними, екологічним та етологічної параметрам. Це дозволяє їм бути буфером між самками і навколишнім середовищем. Через підвищену мінливості під впливом середовища в першу чергу буде елімінована якась частина самців. Ті, що вижили, т. Е. Краще пристосовані до цих впливів, самці, схрещуючи з самками, різко збільшать подібних собі самців, а також зрушать середній генотип самок в сторону найбільшої схожості з собою.
Подібну думку про те, що співвідношення статей визначає швидкість перетворення популяції висловлював також Д. Гізель (Giesel, 1972). Диференційована смертність самців і самок повинна бути непрямим підкріпленням справедливості гіпотези В. А. Геодакяна, і багато дослідників підтверджують її (Fisher, 1930; Keyffitz, Flieger, 1971; Giesel, 1972; Charnov, 1974) »(кінець цитати - курсив автора).
Тихоокеанські лососі і нерки, зокрема, підтверджує деякі положення, висловлені В. А. Геодакяном (Коновалов, 1980). Так, наприклад, мінливість по таким важливим ознаками, як розміри і маса тіла у нерки, у самців істотно перекриває мінливість самок.
Відбір при такій розмірній структурі буде по-різному впливати на великих (3-4 роки життя в море) або дрібних (частіше 1-2 роки життя в море) самців, що було виявлено при дослідженні виїданням нерки ведмедями. Якщо вважати дрібних самців, які не зуміли залишити потомство через ассортативного (виборчого) схрещування, характерного для лососів, також елімініруемимі, то смертність самців відразу різко зростає в порівнянні зі смертністю самок (Коновалов, 1980).
Для нерки ассортативное схрещування виражається в тому, що великі самці завжди отримують перевагу у самок, тоді як середні і дрібні самці не допускаються до самкам великими самцями, та й самки відганяють дрібних і середніх самців при наявності великих (Hanson, Smith, 1967; Коновалов, 1980).
Для інших видів тихоокеанських лососів - горбуші, кети і кижуча, також притаманне ассортативное схрещування (Чебанов, 1983). Ймовірно, воно має місце і у чавичі і у Сіми, т. К. Всі ці риби належать до одного роду Oncorhynchus.
Таким чином, статеве розмноження може забезпечити володіють їм види низку переваг в тому випадку, коли необхідний досконалий адаптаційний механізм до впливів навколишнього середовища. Складні життєві цикли тварин, жорсткі вимоги до гранично точної і швидкої передачі інформації про результати взаємодії популяції із середовищем дають перевагу статевого розмноження. Серед риб тихоокеанські лососі відносяться до видів, які мають складний життєвий цикл. Перед лососями поставлені вкрай жорсткі вимоги до прийнятно-оперативної передачі інформації від покоління до покоління. І не виключено, що у тихоокеанських лососів існує досконалий механізм визначення статі і формування статевої структури (Коновалов, 1980).
література
Бугаєв В.Ф. 2007. Риби басейну річки Камчатки (чисельність, промисел, проблеми) // Петропавловськ-Камчатський: Видавництво "Камчатпресс". - 192 с.
Токранов Василь Вікторович А.М., В.Ф. Бугаєв. 2011. Де крупніше лососі ?: довідковий посібник // Петропавловськ-Камчатський: Изд-во "Камчатпресс" ". - 72 с.
На фото: гермафродит нерки з р. Бушуєвої (басейн оз. Азабачье). Такі риби попадаються один раз на 5-7 тисяч нормальних особин (по: Бугайов, 2007).
рецензії
Привіт, Віктор!У популяціях сріблястого карася немає самців. Ікра самок цього карася, скажімо так, активується молоками інших риб. При цьому схрещування не відбувається, і ДНК сріблястого карася залишається постійною.
Якщо послухати Дарвіна, то такий карась приречений в постійно мінливих умовах середовища проживання. Однак на Челябінських озерах сріблястий карась переважає і, як кажуть місцеві рибалки, витісняє золотистого. Можу навести приклад Щелкунского озера, в якому золотистого карася вже не залишилося, а сріблястий клює незрівнянно.
Це я все до того, що біологічному виду важлива не тільки мінливість, але і сталість ідеально пристосованих особин. Золотистий карась, як виявилося, схильний до виродження.
З повагою,
Віктор.
Віктор Бабінцев 02.12.2013 10:26 • Заявити про порушення Віктор, спеціально для тебе я поставив статтю про срібного карася. Сподіваюся, що в ній знайдеш деякі відповіді на питання, що цікавлять тебе питання.
Вже давно я посилав тобі е-мейл вийти на більш тісний зв'язок, але ти не відповів.
З повагою, Віктор
Віктор Бугаєв 06.12.2013 2:39 Заявити про порушення Привіт, Віктор!
Дуже дякую!
За електронну адресу прошу вибачення. У мене два компа і обидва останнім часом постійно то глючат, то взагалі в ремонті, то мені самому не до них.
Віктор Бабінцев 06.12.2013 5:03 Заявити про порушення 2011. Де крупніше лососі ?


